





![]()
![]()
![]()
 Por Alane Beatriz Vermelho
Por Alane Beatriz Vermelho
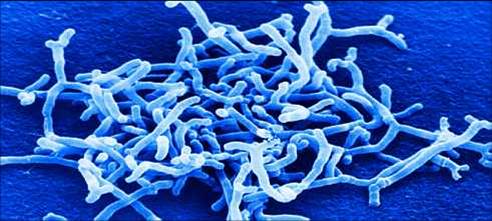
Na última edição do periódico “Nature reviews Microbiology”, foi publicada uma interessante revisão sobre a relação entre o metabolismo dos carboidratos com os micro-organismos da microbiota intestinal. Glicanas e polissacarídeos provenientes de alimentos vegetais (amido, hemicelulose e pectina), cartilagem e tecidos animais (glicosmanoglicanas e N- glicanas) e o muco (glicanas O- Ligadas) trazem uma enorme variedade de moléculas para o ambiente dos micro-organismos intestinais. Como exemplo, podemos citar a glicoproteína mucina do muco, que pode ter centenas de diferentes estruturas ligadas a glicoproteína.
A microbiota que se estabelece no intestino após o nascimento tem um efeito profundo na saúde e fisiologia humana, beneficiando a modulação do desenvolvimento imunológico, auxiliando na digestão de nutrientes, sintetizando vitaminas e inibindo a colonização de patógenos. Anormalidades nessa microbiota, denomiada de disbiose, pode levar a sérios problemas como doenças inflamatórias intestinais, câncer de colon, colite associada a antibiótico e obesidade. A disbiose é caracterizada por um desequilíbrio nos micro-organismos intestinais que causa uma proliferação de outros que são prejudiciais a saúde. Pode ocasionar também um aumento do fluxo de vias metabólicas prejudiciais.
Esse tema tem sido alvo de intensos estudos nos últimos anos e hoje se sabe que um dos fatores principais que regulam a composição e fisiologia dessa microbiota no intestino são os carboidratos presentes no intestino e provenientes da dieta ou do muco intestinal. Nesse panorama, se observa a diversidade de preferência por determinado carboidratos entre os micro-organismos residentes. Muitas dessas glicanas e polissacarídeos não podem ser hidrolisados pelas enzimas codificadas pelo genoma humano e, desse modo, a fermentação microbiana tem um papel fundamental na transformação dessas glicanas não digeríveis em ácidos graxos de cadeia curta (SCFAs) como, por exemplo, os ácidos propriônico, acético e butírico. Esses ácidos servem como nutrientes para os colonócitos e outras células epiteliais do intestino.
No início da colonização da microbiota, os carboidratos disponíveis são provenientes do leite materno cujos componentes primários são a lactose, glactose, N-acetilglicosamina, fucose, acido siálico e uma mistura de oligossacarídeos complexos (HMOs). Esses oligossacarídeos são glicanas com grande diversidade compostas de lactose com ramificações ou N- acetil lactosmamina, com ácidos siálicos e fucose em suas cadeias. A microbiota nascente tem basicamente 4 filos de micro-organimos: Bacteroidetes, Proteobacteria, Firmicutes e Actinobacteria. Em crianças alimentadas exclusivamente com o leite materno, predominam os gêneros Lactobacillus spp. e Bifidobacterium spp. que metabolizam muito bem o oligossacarídeos (HMOs). Crianças alimentadas com leites formulados têm menos Lactobacillus e Bifidobacterium spp e um aumento significativo de Clostridium spp., Bacteroides spp. e de membros da família Enterobacteriaceae. Ao serem introduzidos na alimentação, cereais, frutas e vegetais outros micro-organismos aparecem como bactérias Gram-negativas como os Bacterióides, espécies do filo Firmicutes e as Actinobactérias. Nas bactérias dos adultos predominam os filos Firmicutes e Bacteroidetes.
O tipo de alimentação é um fator importante. Estudos feitos com crianças africanas, que têm uma dieta rica em fibras e crianças européias com uma alimentação baixa em fibras, mostraram diferenças significativas. No primeiro grupo, a microbiota mostrou ser formada mais por espécies de Bacteroidetes e Actinobacterias do que por Firmicutes e proteobacteria. O oposto foi observado nas crianças européias. Diferenças também foram observadas nos gêneros encontrados: nas crianças africanas predominam o Prevotella spp. e Xylanibacter spp., enquanto nas européias, os gênerosBacteroides spp. e Alistipes spp prevalecem dentro dos Bacteroidetes.
Vários fatores afetam a utilização das glicanas pelas bactérias intestinais. A degradação destas glicanas pode ser dificultada porque muitos destes substratos estão em microambientes como a camada de mucosa do intestino. Outras estão em alimentos que possuem glicanas como celuloses, hemicelulose e pectina na parede celular destas plantas. O amido pode estar em grânulos insolúveis resistentes às enzimas degradativas. Cozimento, moagem e outras metodologias usadas na preparação dos alimentos podem minimizar estes problemas, mas são as enzimas microbianas que conseguem degradar estas glicanas. Algumas espécies, como o endosimbionte Bacteroides thetaiotaomicron, por exemplo, pode degradar mais de 12 tipo de glicanas.
Estratégias diferentes para degradar os carboidratos são desenvolvidas por esses micro-organismos em relação à organização e expressão dos genes, tipo e números de enzimas envolvidas e mecanismo de transporte. No intestino, existem os microambientes diferentes, passando pelo íleo e colon até o reto, com tecidos com características histológicas peculiares em relação à espessura da camada mucosa que vão alterar a disponibilidade das glicanas.
Suplementações com fibras prebióticas da dieta como a inulina (β- frutana de cadeia longa) e fruto-oligossacarideos têm sido eficientes para minimizar os efeitos de uma dieta rica em gordura. Essas fibras promovem o crescimento do Bifidobacterium spp., Roseburia spp. e Faecalibacterium prausnitzii. As fibras previnem o câncer de colo em ratos e reduz inflamações. Recentemente, formulações contendo arabinoxialanas e chitina-glicana foram promissoras em restaurar o balaço Bacterioides/ Firmicutes na comunidade microbiana, melhorando as funções, mesmo em dietas ricas em gorduras. Os prebióticos ajudam a reduzir o colesterol porque estimulam a microbiota a produzir propianato que é transportado pela corrente sanguínea até o fígado, inibindo a síntese de colesterol. A adição de outros micro-organismos para compor a microbiota, como os probióticos, também é de grande utilidade.
Todos esses estudos, muito bem abordados nesta revisão, mostraram que a disponibilidade das glicanas e polissacarídeos é um dos principais fatores na determinação da relação dos carboidratos com a microbiota intestinal.
Fonte: Nature Reviews Microbiology
 Por Raquel Regina Bonelli
Por Raquel Regina Bonelli
Historicamente consideramos a resistência a antimicrobianos apenas no contexto clínico, focando no enorme potencial de disseminação de microrganismos resistentes que o uso indiscriminado de antibióticos provoca. De fato, os altos índices de resistência observados em bactérias patogênicas são um dos maiores desafios da microbiologia médica para o século XXI.
Observando o desenvolvimento histórico da resistência aos diferentes antibióticos, fica claro que a incorporação de uma nova droga no plantel de medicamentos disponíveis para o tratamento de doenças infecciosas logo é seguida pela emergência de cepas resistentes. Um dos mecanismos que pode levar a isso é a ocorrência de mutações pontuais do DNA da bactéria, que, ao acaso, podem conferir uma vantagem e assim possibilitar a seleção da cepa resistente, assim que essa entrar em contato com o antibiótico.
No entanto, a resistência observada para a maior parte dos antibióticos não se estabelece dessa forma, sendo, ao contrário, incorporada pronta a partir de outras bactérias, através de elementos genéticos móveis. Pesquisas com o objetivo de identificar a origem desses genes apontam que uma possível fonte são bactérias produtoras de antibióticos na natureza, o que é bastante racional, uma vez que estas necessitam de mecanismos para se proteger do próprio antimicrobiano que produzem. Mas, surpreendentemente, muitos genes de resistência são também oriundos de bactérias que não produzem, nem tiveram contato com concentrações potencialmente ativas de antibióticos. Esse cenário é intrigante porque levanta questões como: que funções biológicas teriam esses genes nas suas bactérias de origem, se não para protegê-las de antimicrobianos? Quantos outros genes, ainda desconhecidos em bactérias ambientais, podem a qualquer momento emergir codificando um novo mecanismo contra medicamentos antimicrobianos? Que bactérias são estas, que atuam como silenciosos reservatórios de mecanismos de resistência na natureza?
Possivelmente, ferramentas de metagenômica e sequenciamento de alta performance irão nos aproximar dessas respostas nos próximos anos. Essa abordagem nova, que integra microbiologia médica e ambiental, será certamente valiosa para a compreensão da dinâmica do desenvolvimento da resistência aos antimicrobianos em patógenos humanos e do papel do meio ambiente nesse processo.
Fonte: Martinez, J.L. Natural antibiotic resistance and contamination by antibiotic resistance determinants: the two ages in the evolution of resistance to antimicrobials. Front Microbiol. 2012, 3:1-3.
Para ler mais: Allen HK, et al. Call of the wild: antibiotic resistance genes in natural environments. Nat Rev Microbiol. 2010, 8(4):251-9.
Fonte: Martinez, Science. 2008, 321 (5887): 365-7.
 Por Kátia Regina Netto dos Santos
Por Kátia Regina Netto dos Santos
Staphylococcus lugdunensis é uma espécie de Staphylococcus coagulase-negativa (SCN) raramente isolada de infecções humanas, porém pode apresentar um potencial agressor, similar ao de S. aureus, pela presença de fatores de virulência em comum. Essemicrorganismo tem grande habilidade de estabelecer infecção primária, visto que é encontrado praticamente em toda superfície do corpo humano. Sua primeira descrição foi em 1988, na cidade de Lyon, na França, por Freney e colaboradores, e a palavra Lugdunum refere-se a antiga denominação desta cidade.
Seus baixos percentuais de isolamento podem estar ligados ao fato de compartilhar semelhanças fenotípicas com S. aureus, já que muitos laboratórios de microbiologia utilizam apenas a identificação rápida em lâmina com plasma para detectar o fator “clumping”, no passado denominado de coagulase ligada, em que ambas as espécies são positivas. Inclusive, essa espécie pode carrear o cassete mec de resistência a meticilina, fato frequentemente observado em S. aureus, tornando ainda mais difícil a diferenciação, como verificamos recentemente em um caso de bacteremia causado pelo patógeno em um hospital do Rio de Janeiro (Pereira et al., 2011). Contudo, a detecção de S. lugdunensis por uma PCR, recentemente desenvolvida em nosso laboratório e baseada no gene fbl, que codifica uma proteína que se liga ao fibrinogênio e coagula o plasma (fator “clumping”) pode ser uma ferramenta auxiliar na identificação de amostras clínicas (Pereira et al., 2010).
Comparado a outros SCN, as infecções por S. lugdunensis podem ser muito graves. Essa espécie pode causar abscessos cerebrais associados a meningites, sepse, osteomielite e endocardites, dentre outras. Endocardites severas podem levar o paciente ao óbito em até 50% dos casos. Alguns fatores de invasão como esterases, proteases e lipases podem ser produzidos por cepas de S. lugdunensis. Uma hemolisina fenotipicamente similar a d-hemolisina de S. aureus e denominada SLUSH (“Staphylococcus lugdunensis synergistic haemolysin”) age sinergicamente com a b-hemolisina, produzindo uma hemólise completa em agar-sangue.
Quanto à produção de biofilme, em estudo recente desenvolvido por nosso grupo, verificamos em um grupo de amostras de S. lugdunensis a presença dos genes slush e ica (Pereira et al., 2012). Este último codifica a produção de biofilme, considerado como forte estimulador da síntese de prostaglandinas E2, que por sua vez inibem a proliferação de células T. Ainda nesse estudo a maioria das cepas de uma mesma linhagem de PFGE produziu biofilme, cuja composição foi demonstrada como protéica. Um dado despertou o interesse da comunidade científica, onde apenas amostras que se apresentaram como colônias rugosas foram capazes de aderir e invadir células epiteliais de pulmão. Essas mesmas amostras também produziram uma substância tipo surfactante que possivelmente lhes forneceu esta aparência colonial típica e pode estar associada ao potencial de adesão/invasão celular de amostras de S. lugdunensis. Fotografias de colônias lisas e rugosas de S. lugdunensis analisadas em nosso estudo serão utilizadas para ilustrar a capa da edição de Maio 2012, Vol 61 da revista Journal of Medical Microbiology.
Pereira et al., 2010, FEMS Immunol. Med. Microbiol. vol 58
Pereira et al., 2011, Braz. J. Infect. Dis. vol. 15
Pereira et al., 2012, J. Med Microbiol. vol 61









